A segunda lei de Mendel
A segregação independente de dois ou mais pares de genes
|
|
|
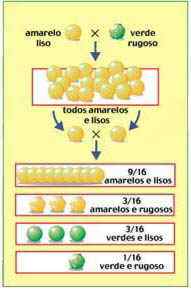
Além de estudar isoladamente diversas características fenotípicas da ervilha, Mendel estudou também a transmissão combinada de duas ou mais características. Em um de seus experimentos, por exemplo, foram considerados simultaneamente a cor da semente, que pode ser amarela ou verde, e a textura da casca da semente, que pode ser lisa ou rugosa.
Plantas originadas de sementes amarelas e lisas, ambos traços dominantes, foram cruzadas com plantas originadas de sementes verdes e rugosas, traços recessivos. Todas as sementes produzidas na geração F1 eram amarelas e lisas.
A geração F2, obtida pela autofecundação das plantas originadas das sementes de F1, era composta por quatro tipos de sementes:
9/16 amarelo-lisas
3/16 amarelo-rugosas
3/16 verde-lisas
1/16 verde-rugosas
|
Em proporções essas frações representam 9 amarelo-lisas: 3 amarelo-rugosas: 3 verde-lisas: 1 verde-rugosa.
Com base nesse e em outros experimentos, Mendel aventou a hipótese de que, na formação dos gametas, os alelos para a cor da semente (Vv) segregam-se independentemente dos alelos que condicionam a forma da semente (Rr). De acordo com isso, um gameta portador do alelo V pode conter tanto o alelo R como o alelo r, com igual chance, e o mesmo ocorre com os gametas portadores do alelo v.
Uma planta duplo-heterozigota VvRr formaria, de acordo com a hipótese da segregação independente, quatro tipos de gameta em igual proporção: 1 VR: 1Vr: 1 vR: 1 vr.
A segunda lei de Mendel
Mendel concluiu que a segregação independente dos fatores para duas ou mais características era um princípio geral, constituindo uma segunda lei da herança. Assim, ele denominou esse princípio segunda lei da herança ou lei da segregação independente, posteriormente chamada segunda lei de Mendel: Os fatores para duas ou mais características segregam-se no híbrido, distribuindo-se independentemente para os gametas, onde se combinam ao acaso.
A proporção 9:3:3:1
Ao estudar a herança simultânea de diversos pares de características. Mendel sempre observou, em F2, a proporção fenotípica 9:3:3:1, conseqüência da segregação independente ocorrida no duplo-heterozigoto, que origina quatro tipos de gameta.
Segregação independente de 3 pares de alelos
Ao estudar 3 pares de características simultaneamente, Mendel verificou que a distribuição dos tipos de indivíduos em F2 seguia a proporção de 27: 9: 9: 9: 3: 3: 3: 1. Isso indica que os genes para as 3 características consideradas segregam-se independentemente nos indivíduos F1, originando 8 tipos de gametas.
Em um dos seus experimentos, Mendel considerou simultaneamente a cor (amarela ou verde), a textura da casca (lisa ou rugosa) e a cor da casca da semente (cinza ou branca).
O cruzamento entre uma planta originada de semente homozigota dominante para as três características (amarelo-liso-cinza) e uma planta originada de semente com traços recessivos (verde-rugosa-branca) produz apenas ervilhas com fenótipo dominante, amarelas, lisas e cinza. Esses indivíduos são heterozigotos para os três pares de genes (VvRrBb). A segregação independente desses três pares de alelos, nas plantas da geração F1, leva à formação de 8 tipos de gametas.
|
|
|
Os gametas produzidos pelas plantas F1 se combinam de 64 maneiras possíveis (8 tipos maternos X 8 tipos paternos), originando 8 tipos de fenótipos.
Determinando o número de tipos de gametas na segregação independente
Para determinar o número de tipos de gametas formados por um indivíduo, segundo a segregação independente, basta aplicar a expressão 2n, em que n representa o número de pares de alelos no genótipo que se encontram na condição heterozigota.
Obtendo a Proporção 9:3:3:1 sem Utilizar o Quadro de Cruzamentos
Genótipo
|
Valor de n
|
2n
|
Número de gametas
|
AA
|
0
|
20
|
1
|
Aa
|
1
|
21
|
2
|
AaBB
|
1
|
21
|
2
|
AaBb
|
2
|
22
|
4
|
AABbCCDd
|
2
|
22
|
4
|
AABbCcDd
|
3
|
23
|
8
|
AaBbCcDd
|
4
|
24
|
16
|
AaBbCcDdEe
|
5
|
25
|
32
|
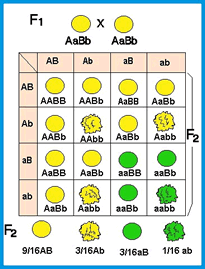
A 2º lei de Mendel é um exemplo de aplicação direta da regra do E de probabilidade, permitindo chegar aos mesmos resultados sem a construção trabalhosa de quadro de cruzamentos. Vamos exemplificar, partindo do cruzamento entre suas plantas de ervilha duplo heterozigotas:
P: VvRr X VvRr
- Consideremos, primeiro, o resultado do cruzamento das duas características isoladamente:
Vv X Vv
|
Rr X Rr
|
3/4 sementes amarelas
|
3/4 sementes lisas
|
1/4 sementes verdes
|
1/4 sementes rugosas
|
- Como desejamos considerar as duas características simultaneamente, vamos calcular a probabilidade de obtermos sementes amarelas e lisas, já que se trata de eventos independentes. Assim,
sementes amarelas E sementes lisas
|
3/4 X 3/4 = 9/16
|
- E a probabilidade de obtermos sementes amarelas e rugosas:
sementes amarelas E sementes rugosas
|
3/4 X 1/4 = 3/16
|
- Agora a probabilidade de obtermos sementes verdes e lisas:
sementes verdes E sementes lisas
|
1/4 X 3/4 = 3/16
|
- Finalmente, a probabilidade de nós obtermos sementes verdes e rugosas:
sementes verdes E sementes rugosas
|
1/4 X 1/4 = 1/16
|
Utilizando a regra do E, chegamos ao mesmo resultado obtido na construção do quadro de cruzamentos com a vantagem da rapidez na obtenção da resposta.
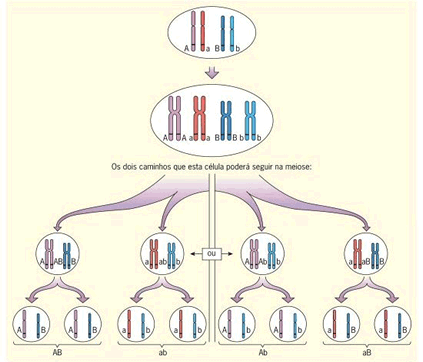
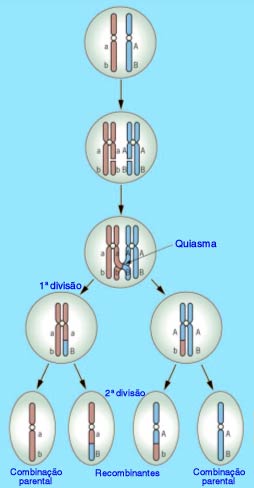
A relação Meiose e 2ª Lei de Mendel
Existe uma correspondência entre as leias de Mendel e a meiose. Acompanhe na figura o processo de formação de gametas de uma célula de indivíduo diíbrido, relacionando-o à 2ª Lei de Mendel. Note que, durante a meiose, os homólogos se alinham em metáfase e sua separação ocorre ao acaso, em duas possibilidades igualmente viáveis. A segregação independente dos homólogos e, consequentemente, dos fatores (genes) que carregam, resulta nos genótipos AB, ab, Ab e aB.
A 2ª Lei de Mendel é sempre obedecida?
A descoberta de que os genes estão situados nos cromossomos gerou um impasse no entendimento da 2º Lei de Mendel. Como vimos, segundo essa lei, dois ou mais genes não-alelos segregam-se independentemente, desde que estejam localizados em cromossomos diferentes. Surge, no entanto, um problema. Mendel afirmava que os genes relacionados a duas ou mais características sempre apresentavam segregação independente. Se essa premissa fosse verdadeira, então haveria um cromossomo para cada gene. Se considerarmos que existe uma infinidade de genes, haveria, então, uma quantidade assombrosa de cromossomos, dentro de uma célula, o que não é verdade. Logo, como existem relativamente poucos cromossomos no núcleo das células e inúmeros genes, é intuitivo concluir que, em cada cromossomo, existe uma infinidade de genes, responsáveis pelas inúmeras características típicas de cada espécie. Dizemos que esses genes presentes em um mesmo cromossomo estão ligados ou em linkage e caminham juntos para a formação dos gametas.
Assim a 2ª lei de Mendel nem sempre é obedecida, bastando para isso que os genes estejam localizados no mesmo cromossomo, ou seja, estejam em linkage.
Linkage
Genes unidos no mesmo cromossomo
|
|
|
T. H. Morgan e seus colaboradores trabalharam com a mosca da fruta, Drosophila melanogaster, e realizaram cruzamentos em que estudaram dois ou mais pares de genes, verificando que, realmente, nem sempre a 2ª Lei de Mendel era obedecida. Concluíram que esses genes não estavam em cromossomos diferente, mas, sim, encontravam-se no mesmo cromossomo (em linkage).
|
Um dos cruzamentos efetuados por Morgan
Em um dos seus experimentos, Morgan cruzou moscas selvagens de corpo cinza e asas longas com mutantes de corpo preto e asas curtas (chamadas de asas vestigiais). Todos os descendentes de F1apresentavam corpo cinza e asas longas, atestando que o gene que condiciona corpo cinza (P) domina o que determina corpo preto (p), assim como o gene para asas longas (V) é dominante sobre o (v) que condiciona surgimento de asas vestigiais.
A seguir Morgan cruzou descendentes de F1 com duplo-recessivos (ou seja, realizou cruzamentos testes). Para Morgan, os resultados dos cruzamentos-teste revelariam se os genes estavam localizados em cromossomos diferentes (segregação-independente) ou em um mesmo cromossomo (linkage).
Surpreendentemente, porém, nenhum dos resultados esperados foi obtido. A separação e a contagem dos decendentes de F2 revelou o seguinte resultado:
- 41,5% de moscas com o corpo cinza e asas longas;
- 41,5% de moscas com o corpo preto e asas vestigiais;
- 8,5% de moscas com o corpo preto e asas longas;
- 8,5% de moscas com o corpo cinza e asas vestigiais.
Ao analisar esse resultado, Morgan convenceu-se de que os genes P e V localizavam-se no mesmo cromossomo. Se estivessem localizados em cromossomos diferentes, a proporção esperada seria outra (1: 1: 1: 1). No entanto, restava a dúvida: como explicar a ocorrência dos fenótipos corpo cinza/asas vestigiais e corpo preto/asas longas?
A resposta não foi difícil de ser obtida. Por essa época já estava razoavelmente esclarecido o processo da meiose. Em 1909, o citologista F. A. Janssens (1863-1964) descreveu o fenômeno cromossômico conhecido como permutação ou crossing over, que ocorre durante a prófase I da meiose e consiste na troca de fragmentos entre cromossomos homólogos.
Em 1911, Morgan usou essa observação para concluir que os fenótipos corpo cinza/asas vestigiais e corpo preto/asas longas eram recombinantes e devido a ocorrência de crossing-over.
Como diferenciar Segregação independente (2ª Lei de Mendel) de Linkage?
Quando comparamos o comportamento de pares de genes para duas características para a segunda lei de Mendel com a ocorrência de linkage e crossing-over em um cruzamento genérico do tipo AaBb X aabb, verificamos que em todos os casos resultam quatro fenótipos diferentes:
- Dominante/dominante
- Dominante/recessivo
- Recessivo/dominante
- Recessivo/recessivo.
A diferença em cada caso está nas proporções obtidas. No caso da 2ª lei de Mendel, haverá 25% de cada fenótipo. No linkage com crossing, todavia, os dois fenótipos parentais surgirão com frequência maior do que as frequências dos recombinantes.
A explicação para isso reside no fato de, durante a meiose a permuta não ocorrer em todas as células, sendo, na verdade, um evento relativamente raro. Por isso, nos cruzamentos PpVv X ppvv, da pagina anterior, foram obtidos 83% de indivíduos do tipo parental (sem crossing) e 17% do tipo recombinantes (resultantes da ocorrência de permuta).
Frequentemente, nos vários cruzamentos realizados do tipo AaBb X aabb, Morgan obteve os dois fenótipos parentais (AaBb e aabb), na proporção de 50% cada. Para explicar esse resultado, ele sugeriu a hipótese que os genes ligados ficam tão próximos um do outro que dificultam a ocorrência de crossing over entre eles. Assim, por exemplo, o gene que determina a cor preta do corpo da drosófila e o gene que condiciona a cor púrpura dos olhos ficam tão próximos que entre eles não ocorre permuta. Nesse caso se fizermos um cruzamento teste entre o duplo-heterozidoto e o duplo-recessivo, teremos nos descendentes apenas dois tipos de fenótipos, que serão correspondentes aos tipos parentais.
Os arranjos “cis” e “trans” dos genes ligados
|
|
|
Considerando dois pares de genes ligados, como, por exemplo, A/a e B/b, um indivíduo duplo heterozigoto pode ter os alelos arranjados de duas maneiras nos cromossomos:
Os alelos dominantes A e B se situam em um cromossomo, enquanto os alelos recessivos a e b se situam no homólogo correspondente. Esse tipo de arranjo é chamado de Cis. O alelo dominante A e o alelo recessivo b se situam em um cromossomo, enquanto o alelo recessivo a e o alelo dominante B, se situam no homólogo correspondente. Esse tipo de arranjo é chamado de Trans.
|
Podemos descrever esses arranjos, usando um traço duplo ou simples para descrever o cromossomo, ou mais simplificadamente, o arranjo pode ser descrito como AB/ab para Cis e Ab/aB para trans. O arranjo cis e trans dos alelos no duplo-heterozigoto pode ser facilmente identificado em um cruzamento teste. No caso dos machos de Drosófila, se o arranjo for cis (PV/pv), o duplo heterozigoto forma 50% de gametas PV e 50% de gametas pv. Se o arranjo for trans (Pv/pV), o duplo heterozigoto forma 50% de gametas Pv e 50% de pV.
Nas fêmeas de Drosófila, nas quais ocorrem permutações, o arranjo cis ou trans pode ser identificado pela frequência das classes de gametas. As classes mais frequentes indicam as combinações parentais e as menos frequentes as recombinantes.
Herança de grupos sanguíneos
Sistema ABO
Indivíduos podem ter sangue do grupo A, B, AB ou O, dependendo da presença de determinados antígenos nos glóbulos vermelhos. Indivíduos com sangue do tipo A possuem o aglutinogênio A; o B, aglutinogênio B; o AB, os dois antígenos citados, e o O, nenhum.
O plasma sanguíneo, por sua vez, pode abrigar outras duas proteínas denominadas aglutininas anti-A e aglutininas anti-B e são elas as responsáveis pelos problemas decorrentes em transfusões de sangue que não observam a compatibilidade sanguínea. Indivíduos A possuem aglutininas anti-B; indivíduos B, anti-A; indivíduos de sangue tipo O possuem as duas aglutininas e os AB, nenhuma.
Ficou confuso? Observe o quadro:
*IA e IB são codominantes
E, desta forma, a relação de dominância entre estes três alelos é: IA = IB > i.
Observando o esquema, é compreensível o porquê de indivíduos de sangue A recebendo sangue de indivíduo do tipo B (ou vice-versa) podem ter complicações sérias e o porquê de indivíduos do tipo AB serem considerados receptores universais. Da mesma forma, indivíduos do tipo O são considerados doadores universais (com algumas ressalvas), pela ausência de aglutinogênio.
Sistema MN
Em 1927, Landsteiner e Levine descobriram dois aglutinogênios nas hemácias humanas, que foram denominados M e N. Eles verificaram que algumas pessoas apresentavam um desses antígenos, enquanto outras apresentavam os dois juntos. Assim, consideraram três fenótipos: grupo M, grupo N e grupo MN, determinados por um par de alelos, sem relação de dominância:
gene LM (ou M) - condiciona a produção do antígeno M;
gene LN (ou N)- condiciona a produção do antígeno N.
Os anticorpos anti-M e anti-N são produzidos apenas quando o indivíduo de um grupo recebe sangue de indivíduo do outro grupo e, assim sendo, problemas decorrentes da incompatibilidade dos grupos ocorrem apenas quando tal procedimento é feito diversas vezes.
Fator Rh
O sistema Rh é controlado por genes independentes dos genes do sistema ABO. Neste, considera-se um par de genes alelos: “R” e “r”, sendo o primeiro o dominante e com presença de antígeno e o segundo, recessivo e sem antígeno. Indivíduos com antígeno são pertencentes ao grupo Rh+ e os não portadores, do grupo Rh-.
Em 1940, Landsteiner e Wiener descobriram este sistema a partir do sangue do macaco Rhesus (Macaca mulatta). O sangue deste animal, uma vez injetado em cobaias ou em coelhos, provocava nestes a síntese de anticorpos (aglutininas anti-Rh), que podiam promover a aglutinação do sangue doado.
Os descobridores do fator Rh extraíram de cobaias e coelhos soros contendo aglutininas anti-Rh. Em seguida, misturaram o soro com sangue de pessoas diversas e constataram que, em alguns casos, as hemácias se aglutinavam, indicando a presença do fator Rh no sangue humano: pessoas Rh+. Em outros casos, as hemácias não se aglutinavam, indicando a ausência do fator Rh no sangue: pessoas Rh-.
Indivíduos de Rh- só apresentarão anticorpos se receberem hemácias de Rh+. Ao se fazer transfusão de sangue de um doador Rh+ para um receptor Rh-, poderá não ocorrer aglutinação das hemácias doadas. Entretanto, em uma segunda transfusão de sangue deste tipo, poderá provocar a aglutinação das hemácias doadas em razão do acúmulo de aglutininas, podendo promover a aglutinação das hemácias do doador e causar obstrução dos capilares sanguíneos e, inclusive, a morte.
Síndromes
Pleiotropia (do grego pleio = "muito" e tropo = "mudança") é o nome dado aos múltiplos efeitos de um gene. Acontece quando um único gene controla diversas características do fenótipo que muitas vezes não estão relacionadas.
Exemplo de pleiotropia:
A Síndrome de Marfan, também conhecida como Aracnodactilia, é uma desordem do tecido conjuntivo caracterizada por membros anormalmente longos. A doença também afeta outras estruturas do corpo, incluindo o esqueleto, os pulmões, os olhos, o coração e os vasos sanguíneos.
A pleiotropia, também denominada por efeito pleiotrópico, é um fenômeno genético em que um único gene possui controle sobres às manifestações de várias características.
A base genética na maioria dos casos consiste na mutação de um gene situado no cromossomo 15, a fibrilina, ou gene FBN1, importante componente na formação das fibras elásticas. Sua produção anormal resulta em fibras elásticas anormais produzindo as alterações que caracterizam a síndrome. Estas mutações podem ser encontradas em 90% dos indivíduos que preenchem os critérios de diagnóstico clínico.
Deformidades anatômicas e fisiológicas, características em portadores da síndrome de Marfan, são fatores que provocam distúrbio de ajustamento psicológico com limitações no desenvolvimento emocional e social. A aparência física pouco comum nesses indivíduos freqüentemente leva a alterações no esquema corporal, na auto-estima e, conseqüentemente, no desenvolvimento global da personalidade. As deformidades do corpo são responsáveis por colocar o indivíduo em uma posição de destaque negativo junto ao grupo, causando prejuízos muitas vezes determinantes na vida da pessoa tais como, inibição, sentimentos de menos valia e tendência ao isolamento. Por ser uma doença crônica e peculiar, atingindo precocemente o indivíduo, a síndrome pode provocar danos na área afetivo-relacional em cada etapa do desenvolvimento.
Fenilcetonúria (PKU - PhenylKetonUria): doença autossômica recessiva caracterizada pelo defeito/ausência da enzima fenilalanina hidroxilase (PAH).
Esta proteína catalisa o processo de conversão (hidroxilização) da fenilalanina em tirosina. A tirosina está envolvida na síntese da melanina, Portanto, bebês com essa doença geralmente possuem pele, cabelo e olhos mais claros do que seus irmãos que não sofrem dessa doença.
A doença é autossômica recessiva e afeta aproximadamente um em cada dez mil indivíduos da população caucasiana. As pessoas com PKU possuem uma mutação no gene da PAH que muda a estrutura da enzima.
Esta doença pode ser detectada logo após o nascimento através de triagem neonatal (conhecida popularmente por teste do pezinho).
.
O tratamento para fenilcetonúria consiste em limitar a ingestão de fenilalaninas da alimentação
São sintomas da doença não tratada: oligofrenia, atraso do desenvolvimento psicomotor (andar ou falar), convulsões, hiperatividade, tremor e microcefalia. Identificam-se as alterações com cerca de um ano de vida. Praticamente todos os pacientes não tratados apresentam um QI inferior a 50.
O tratamento consiste em uma dieta pobre em fenilalanina (300 a 500 mg/dia em uma criança de 10 anos). A expectativa de vida sem tratamento é baixa, em torno de 30 anos.
Outros sintomas podem incluir:
Atraso mental e das habilidades sociais
Tamanho da cabeça significantemente menor do normal
Hiperatividade
Movimentos incontroláveis de braços e pernas
Retardo mental
Convulsões
Erupções cutâneas
Tremores
Posicionamento incomum das mãos
Se a doença não for tratada ou se os alimentos contendo fenilalanina não forem evitados, um odor "de rato" poderá ser sentido no hálito, na pele e na urina. Esse odor incomum deve-se ao aumento de substâncias de fenilalanina no organismo.
INTERAÇÕES GÊNICAS → Dois ou mais genes interagem entre si par determinar uma única característica
Existem casos em que os alelos de um gene inibem a ação dos alelos de um outro par, que pode ou não estar no mesmo cromossomo. Esse fenômeno é chamado epistasia (do grego epi, sobre, e stasis, parada, inibição). O gene que exerce a ação inibitória é chamado epistático, e o que sofre a inibição é chamado hipostático.
A pelagem no cão labrador é de terminada por epistasia recessiva, onde o alelo E determina o pigmento preto, ee determina o pigmento marrom, B determina a deposição de pigmentos, bb não determina a deposição do pigmento, originando a coloração dourada ou caramelo.
.jpg)
.jpg)
